Medisinsk ekspert av artikkelen

Nye publikasjoner
Metodikk for elektroencefalografi
Sist anmeldt: 04.07.2025

Alt iLive-innhold blir gjennomgått med medisin eller faktisk kontrollert for å sikre så mye faktuell nøyaktighet som mulig.
Vi har strenge retningslinjer for innkjøp og kun kobling til anerkjente medieområder, akademiske forskningsinstitusjoner og, når det er mulig, medisinsk peer-evaluerte studier. Merk at tallene i parenteser ([1], [2], etc.) er klikkbare koblinger til disse studiene.
Hvis du føler at noe av innholdet vårt er unøyaktig, utdatert eller ellers tvilsomt, velg det og trykk Ctrl + Enter.
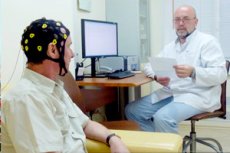
I vanlig praksis registreres EEG ved hjelp av elektroder plassert på intakt hodebunn. Elektriske potensialer forsterkes og registreres. Elektroencefalografer har 16–24 eller flere identiske forsterknings- og registreringsenheter (kanaler) som tillater samtidig registrering av elektrisk aktivitet fra det tilsvarende antallet par elektroder installert på pasientens hode. Moderne elektroencefalografer er databaserte. Forsterkede potensialer konverteres til digital form; kontinuerlig EEG-registrering vises på en skjerm og registreres samtidig på en disk. Etter behandling kan EEG-en skrives ut på papir.
Elektroder som leder potensialer er metallplater eller stenger i forskjellige former med en kontaktflatediameter på 0,5–1 cm. Elektriske potensialer mates til inngangsboksen på elektroencefalografen, som har 20–40 eller flere nummererte kontaktuttak, og ved hjelp av disse kan det tilsvarende antallet elektroder kobles til enheten. I moderne elektroencefalografer kombinerer inngangsboksen en elektrodebryter, en forsterker og en EEG analog-til-digital-omformer. Fra inngangsboksen mates det konverterte EEG-signalet til en datamaskin, ved hjelp av hvilken enhetens funksjoner styres, og EEG registreres og behandles.
EEG registrerer potensialforskjellen mellom to punkter på hodet. Følgelig mates spenninger avledet fra to elektroder til hver kanal på elektroencefalografen: én til "inngang 1" og den andre til "inngang 2" på forsterkningskanalen. En flerkontakts EEG-ledningsbryter lar deg kommutere elektrodene for hver kanal i ønsket kombinasjon. For eksempel, ved å stille inn korrespondansen til den occipitale elektroden til soklen på inngangsboksen "1" på en hvilken som helst kanal, og den temporale elektroden til soklen på boksen "5", kan du dermed registrere potensialforskjellen mellom de tilsvarende elektrodene i denne kanalen. Før arbeidet starter, skriver forskeren flere ledningsdiagrammer ved hjelp av passende programmer, som brukes til å analysere de innhentede registreringene. For å stille inn båndbredden til forsterkeren brukes analoge og digitale høy- og lavfrekvensfiltre. Standardbåndbredden ved EEG-opptak er 0,5-70 Hz.
Elektroencefalogramopptak og -opptak
Opptakselektrodene er plassert slik at alle hoveddelene av hjernen, betegnet med de første bokstavene i deres latinske navn, er representert i flerkanalsopptaket. I klinisk praksis brukes to hoved-EEG-ledningssystemer: det internasjonale 10-20-systemet og et modifisert skjema med et redusert antall elektroder. Hvis det er nødvendig å få et mer detaljert EEG-bilde, er 10-20-skjemaet å foretrekke.
En referanseledning er en der potensialet fra en elektrode plassert over hjernen mates til "inngang 1" på forsterkeren, og fra en elektrode plassert langt fra hjernen til "inngang 2". Elektroden plassert over hjernen kalles oftest aktiv. Elektroden langt fra hjernevevet kalles referanse. Venstre (A1 ) og høyre (A2 ) øreflipper brukes som referanseelektroder. Den aktive elektroden er koblet til "inngang 1" på forsterkeren, og å mate den med et negativt potensialskift får opptakspennen til å avbøyes oppover. Referanseelektroden er koblet til "inngang 2". I noen tilfeller brukes en ledning fra to elektroder (AA) kortsluttet sammen og plassert på øreflippene som referanseelektrode. Siden EEG registrerer potensialforskjellen mellom to elektroder, vil posisjonen til punktet på kurven være likt, men i motsatt retning, påvirket av endringer i potensialet under hvert av elektrodeparene. I referanseledningen genereres et vekslende potensial for hjernen under den aktive elektroden. Under referanseelektroden, som er plassert langt fra hjernen, er det et konstant potensial som ikke går inn i vekselstrømsforsterkeren og ikke påvirker opptaksmønsteret. Potensialforskjellen reflekterer uten forvrengning svingningene i det elektriske potensialet som genereres av hjernen under den aktive elektroden. Imidlertid er området av hodet mellom den aktive elektroden og referanseelektroden en del av den elektriske kretsen "forsterker-objekt", og tilstedeværelsen av en tilstrekkelig intens potensialkilde i dette området, plassert asymmetrisk i forhold til elektrodene, vil påvirke avlesningene betydelig. Følgelig er vurderingen av lokaliseringen av potensialkilden med referanseledningen ikke helt pålitelig.
Bipolar er navnet på ledningen der elektroder plassert over hjernen er koblet til forsterkerens "inngang 1" og "inngang 2". Plasseringen av EEG-opptakspunktet på skjermen påvirkes likt av potensialene under hvert av elektrodeparene, og den registrerte kurven gjenspeiler potensialforskjellen til hver av elektrodene. Derfor er det umulig å bedømme formen på oscillasjonen under hver av dem basert på én bipolar ledning. Samtidig lar analysen av EEG registrert fra flere par elektroder i forskjellige kombinasjoner oss bestemme lokaliseringen av potensialkildene som utgjør komponentene i den komplekse sammendragskurven oppnådd med bipolar ledning.
Hvis det for eksempel finnes en lokal kilde til langsomme oscillasjoner i den bakre temporale regionen, vil tilkobling av de fremre og bakre temporale elektrodene (Ta, Tr) til forsterkerterminalene produsere en registrering som inneholder en langsom komponent som tilsvarer den langsomme aktiviteten i den bakre temporale regionen (Tr), med raskere oscillasjoner generert av den normale hjernemassen i den fremre temporale regionen (Ta) lagt oppå den. For å avklare spørsmålet om hvilken elektrode som registrerer denne langsomme komponenten, kobles elektrodepar på to ekstra kanaler, hvor den ene er representert av en elektrode fra det opprinnelige paret, dvs. Ta eller Tr, og den andre tilsvarer en ikke-temporal avledning, for eksempel F og O.
Det er tydelig at i det nydannede paret (Tr-O), inkludert den bakre temporale elektroden Tr, som er plassert over den patologisk endrede hjernemassen, vil den langsomme komponenten igjen være tilstede. I paret, til hvis innganger aktiviteten fra to elektroder plassert over den relativt intakte hjernen (Ta-F) mates, vil et normalt EEG bli registrert. I tilfelle av et lokalt patologisk kortikalt fokus, fører det å koble elektroden plassert over dette fokuset i et par med et hvilket som helst annet til forekomsten av en patologisk komponent på de tilsvarende EEG-kanalene. Dette lar oss bestemme lokaliseringen av kilden til patologiske svingninger.
Et ytterligere kriterium for å bestemme lokaliseringen av kilden til potensialet av interesse på EEG er fenomenet med oscillasjonsfaseforvrengning. Hvis vi kobler tre elektroder til inngangene på to kanaler på en elektroencefalograf som følger: elektrode 1 til "inngang 1", elektrode 3 til "inngang 2" på forsterker B, og elektrode 2 samtidig til "inngang 2" på forsterker A og "inngang 1" på forsterker B; antar vi at det under elektrode 2 er et positivt skifte i det elektriske potensialet i forhold til potensialet til de resterende delene av hjernen (indikert med "+"-tegnet), er det åpenbart at den elektriske strømmen forårsaket av dette potensialskiftet vil ha motsatt retning i kretsene til forsterkerne A og B, noe som vil reflekteres i motsatt rettede forskyvninger i potensialforskjellen - motfaser - på de tilsvarende EEG-registreringene. Dermed vil de elektriske oscillasjonene under elektrode 2 i registreringene på kanalene A og B bli representert av kurver med samme frekvenser, amplituder og former, men motsatt i fase. Når man bytter elektroder over flere kanaler i en elektroencefalograf i form av en kjede, vil antifase-oscillasjoner av potensialet som studeres bli registrert langs de to kanalene hvis motsatte innganger en felles elektrode er koblet til, plassert over kilden til dette potensialet.
 [ 6 ], [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ]
[ 6 ], [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ]
Regler for registrering av elektroencefalogrammer og funksjonstester
Under undersøkelsen skal pasienten være i et lys- og lydtett rom i en komfortabel stol med lukkede øyne. Personen observeres direkte eller med videokamera. Under opptaket markeres viktige hendelser og funksjonstester med tusjer.
Når man tester åpning og lukking av øynene, vises karakteristiske elektrookulogram-artefakter på EEG-et. De resulterende EEG-endringene lar oss identifisere graden av kontakt hos individet, bevissthetsnivået og grovt estimere reaktiviteten til EEG-et.
For å oppdage hjernens respons på ytre påvirkninger brukes enkeltstimuli i form av et kort lysglimt eller et lydsignal. Hos pasienter i komatøs tilstand er det tillatt å bruke nociseptive stimuli ved å trykke en negl mot roten av pasientens pekefinger.
For fotostimulering brukes korte (150 μs) lysglimt nær hvitt i spekteret og med tilstrekkelig høy intensitet (0,1–0,6 J). Fotostimulatorer muliggjør presentasjon av blitzserier som brukes til å studere rytmeassimileringsreaksjonen – evnen til elektroencefalografiske oscillasjoner til å reprodusere rytmen til eksterne stimuli. Normalt uttrykkes rytmeassimileringsreaksjonen godt ved en flimrende frekvens nær EEGs egne rytmer. Rytmiske assimileringsbølger har størst amplitude i occipitalregionene. Ved epileptiske anfall relatert til lysfølsomhet avslører rytmisk fotostimulering en fotoparoksysmal respons – en generalisert utladning av epileptiform aktivitet.
Hyperventilering utføres primært for å indusere epileptiform aktivitet. Personen blir bedt om å puste dypt og rytmisk i 3 minutter. Respirasjonsfrekvensen bør være innenfor 16–20 per minutt. EEG-registrering starter minst 1 minutt før hyperventileringen starter og fortsetter gjennom hele hyperventileringen og i minst 3 minutter etter at den er slutt.