Medisinsk ekspert av artikkelen

Nye publikasjoner
hypofysen
Sist anmeldt: 04.07.2025

Alt iLive-innhold blir gjennomgått med medisin eller faktisk kontrollert for å sikre så mye faktuell nøyaktighet som mulig.
Vi har strenge retningslinjer for innkjøp og kun kobling til anerkjente medieområder, akademiske forskningsinstitusjoner og, når det er mulig, medisinsk peer-evaluerte studier. Merk at tallene i parenteser ([1], [2], etc.) er klikkbare koblinger til disse studiene.
Hvis du føler at noe av innholdet vårt er unøyaktig, utdatert eller ellers tvilsomt, velg det og trykk Ctrl + Enter.
Hypofysen (hypofysen, s.glandula pituitaria) ligger i hypofysens fossa i sella turcica i kilebenet og er atskilt fra kraniehulen ved en utløper i dura mater i hjernen, som danner diafragma i sella. Gjennom åpningen i dette diafragmaet er hypofysen forbundet med infundibulum i hypothalamus i diencephalon. Hypofysens tverrgående størrelse er 10–17 mm, anteroposterior er 5–15 mm, vertikal er 5–10 mm. Hypofysens masse hos menn er omtrent 0,5 g, hos kvinner - 0,6 g. Hypofysen er dekket utvendig av en kapsel.
I samsvar med hypofysens utvikling fra to forskjellige grunnleggere, skilles det mellom to lapper i organet - den fremre og den bakre. Adenohypofysen, eller fremre lapp (adenohypophysis, s.lobus anterior), er større og utgjør 70-80 % av hypofysens totale masse. Den er tettere enn bakre lapp. I fremre lapp skilles det ut en distal del (pars distalis), som opptar den fremre delen av hypofysefossaen, en mellomliggende del (pars intermedia), som ligger på grensen til bakre lapp, og en tuberøs del (pars tuberalis), som går oppover og er forbundet med hypothalamus infundibulum. På grunn av overfloden av blodårer har fremre lapp en blek gul farge med et rødlig skjær. Parenkymet i den fremre hypofysen er representert av flere typer kjertelceller, mellom hvilke sinusformede blodkapillærer er plassert. Halvparten (50 %) av adenohypofysecellene er kromofile adenocytter, som har finkornede granuler i cytoplasmaet som farges godt med kromsalter. Dette er syrefile adenocytter (40 % av alle adenohypofyseceller) og basofile adenocytter (10 %). Basofile adenocytter inkluderer gonadotrope, kortikotrope og tyreotrope endokrinocytter. Kromofobe adenocytter er små, de har en stor kjerne og en liten mengde cytoplasma. Disse cellene regnes som forløpere til kromofile adenocytter. De andre 50 % av adenohypofysecellene er kromofobe adenocytter.

Nevrohypofysen, eller bakre lobb (nevrohypophysis, s.lobus posterior), består av nevrallappen (lobus nervosus), som ligger i den bakre delen av hypofysen, og trakten (infundibulum), som ligger bak den tuberøse delen av adenohypofysen. Bakre lobb av hypofysen dannes av nevrogliaceller (hypofyseceller), nervefibre som går fra de nevrosekretoriske kjernene i hypothalamus til nevrohypofysen, og nevrosekretoriske blodlegemer.
Hypofysen er, via nervefibre (baner) og blodårer, funksjonelt forbundet med hypothalamus i diencephalon, som regulerer aktiviteten til hypofysen. Hypofysen og hypothalamus, sammen med deres nevroendokrine, vaskulære og nerveforbindelser, regnes vanligvis som hypothalamus-hypofysesystemet.
Hormoner i den fremre og bakre hypofysen påvirker mange av kroppens funksjoner, først og fremst gjennom andre endokrine kjertler. I den fremre hypofysen produserer acidofile adenocytter (alfaceller) somatotropisk hormon (veksthormon), som deltar i reguleringen av vekst- og utviklingsprosesser hos unge organismer. Kortikotrope endokrinocytter skiller ut adrenokortikotropisk hormon (ACTH), som stimulerer utskillelsen av steroidhormoner fra binyrene. Tyrotrope endokrinocytter skiller ut tyrotropisk hormon (TSH), som påvirker utviklingen av skjoldbruskkjertelen og aktiverer produksjonen av hormonene. Gonadotrope hormoner: follikkelstimulerende (FSH), luteiniserende (LH) og prolaktin - påvirker kroppens seksuelle modning, regulerer og stimulerer utviklingen av follikler i eggstokken, eggløsning, vekst av melkekjertlene og melkeproduksjon hos kvinner, samt spermatogenese hos menn. Disse hormonene produseres av basofile adenocytter (betaceller). Lipotrope faktorer i hypofysen skilles også ut her, som påvirker mobilisering og utnyttelse av fett i kroppen. I den mellomliggende delen av forlappen dannes melanocyttstimulerende hormon, som kontrollerer dannelsen av pigmenter - melaniner - i kroppen.
Nevrosekretoriske celler i de supraoptiske og paraventrikulære kjernene i hypothalamus produserer vasopressin og oksytocin. Disse hormonene transporteres til cellene i den bakre hypofysen langs aksoner som utgjør hypothalamus-hypofysen. Fra den bakre hypofysen kommer disse stoffene inn i blodet. Hormonet vasopressin har en vasokonstriktiv og antidiuretisk effekt, som det også kalles antidiuretisk hormon (ADH) for. Oksytocin har en stimulerende effekt på kontraktiliteten til livmormusklene, øker melkesekresjonen fra den ammende melkekjertlen, hemmer utviklingen og funksjonen til corpus luteum og påvirker endringer i tonen i den glatte (ikke-stripete) muskulaturen i mage-tarmkanalen.
Utvikling av hypofysen
Hypofysens fremre lobe utvikler seg fra epitelet i ryggveggen i munnhulen i form av en ringformet utvekst (Rathkes pose). Denne ektodermale utveksten vokser mot bunnen av den fremtidige tredje ventrikkelen. Mot den, fra den nedre overflaten av den andre hjernevesikelen (den fremtidige bunnen av den tredje ventrikkelen), vokser en prosess, hvorfra den grå tuberkelen i infundibulum og den bakre loben i hypofysen utvikler seg.
Kar og nerver i hypofysen
Fra de indre halspulsårene og karene i hjernens arteriekrets leder de øvre og nedre hypofysearteriene til hypofysen. De øvre hypofysearteriene går til den grå kjernen og infundibulum i hypothalamus, anastomoser med hverandre her og danner kapillærer som trenger inn i hjernevevet - det primære hemokapillære nettverket. Fra de lange og korte løkkene i dette nettverket dannes portvenene, som leder til hypofysens fremre lop. I parenkymet til hypofysens fremre lop brytes disse venene opp i brede sinusformede kapillærer, og danner det sekundære hemokapillære nettverket. Hypofysens bakre lop forsynes hovedsakelig med blod fra den nedre hypofysearterien. Det er lange arterielle anastomoser mellom de øvre og nedre hypofysearteriene. Utstrømningen av venøst blod fra det sekundære hemokapillære nettverket skjer gjennom et system av vener som strømmer inn i de kavernøse og interkavernøse bihulene i hjernens dura mater.
Sympatiske fibre som kommer inn i organet sammen med arteriene deltar i hypofysens innervering. Postganglioniske sympatiske nervefibre går fra plexus i den indre halspulsåren. I tillegg finnes en rekke avslutninger av prosessene til nevrosekretoriske celler som ligger i kjernene i hypothalamus, i den bakre loben av hypofysen.
Aldersrelaterte egenskaper ved hypofysen
Gjennomsnittsvekten til hypofysen hos nyfødte når 0,12 g. Organets vekt dobles ved 10-årsalderen og tredobles ved 15-årsalderen. Ved 20-årsalderen når hypofysens vekt sitt maksimum (530-560 mg) og forblir nesten uendret i påfølgende aldersperioder. Etter 60 år observeres en liten reduksjon i vekten av denne endokrine kjertelen.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Hypofysehormoner
Enheten av nervøs og hormonell regulering i kroppen sikres av den nære anatomiske og funksjonelle forbindelsen mellom hypofysen og hypothalamus. Dette komplekset bestemmer tilstanden og funksjonen til hele det endokrine systemet.
Den viktigste endokrine kjertelen som produserer en rekke peptidhormoner som direkte regulerer funksjonen til de perifere kjertlene, er hypofysen. Det er en rødgrå bønneformet formasjon dekket med en fibrøs kapsel som veier 0,5-0,6 g. Den varierer noe avhengig av kjønn og alder. Den allment aksepterte inndelingen av hypofysen i to lapper som er forskjellige i utvikling, struktur og funksjon, er fortsatt: den fremre distale - adenohypofysen og den bakre - nevrohypofysen. Den første utgjør omtrent 70 % av kjertelens totale masse og er konvensjonelt delt inn i den distale, infundibulære og mellomliggende delen, den andre - i den bakre delen, eller lappen, og hypofysestilken. Kjertelen ligger i hypofysefossaen i sella turcica i sphenoidbenet og er koblet til hjernen gjennom stilken. Den øvre delen av den fremre lappen er dekket av den optiske chiasmen og de optiske kanalene. Blodtilførselen til hypofysen er svært rikelig og utføres av grener av den indre halspulsåren (hypofysearteriene øvre og nedre), samt av grener av arteriekretsen i storhjernen. De øvre hypofysearteriene deltar i blodtilførselen til adenohypofysen, og de nedre - nevrohypofysen, i kontakt med de nevrosekretoriske endene av aksonene i de store cellekjernene i hypothalamus. De førstnevnte går inn i den mediane eminensen av hypothalamus, hvor de sprer seg i et kapillærnettverk (primær kapillærpleksus). Disse kapillærene (som terminalene til aksonene til de små nevrosekretoriske cellene i den mediobasale hypothalamus er i kontakt med) samles i portalvener som går ned langs hypofysestilken inn i adenohypofysens parenkym, hvor de igjen deler seg i et nettverk av sinusformede kapillærer (sekundær kapillærpleksus). Dermed kommer blodet, som tidligere har passert gjennom hypothalamus mediane eminens, hvor det er beriket med hypothalamiske adenohypofysotrope hormoner (frigjørende hormoner), inn i adenohypofysen.
Utstrømningen av blod mettet med adenohypofysehormoner fra en rekke kapillærer i sekundærpleksen utføres gjennom venesystemet, som igjen strømmer inn i venøse bihuler i dura mater og deretter inn i den generelle blodbanen. Dermed er hypofysens portalsystem med en synkende retning av blodstrømmen fra hypothalamus en morfofunksjonell komponent i den komplekse mekanismen for nevrohumoral kontroll av adenohypofysens tropiske funksjoner.
Hypofysen er innervert av sympatiske fibre som følger hypofysearteriene. De stammer fra postganglioniske fibre som går gjennom den indre carotis plexus, koblet til de øvre cervikale gangliene. Det er ingen direkte innervering av adenohypofysen fra hypothalamus. Den bakre loben mottar nervefibre fra de nevrosekretoriske kjernene i hypothalamus.
Adenohypofysen er en svært kompleks formasjon i sin histologiske arkitektur. Den har to typer kjertelceller - kromofobe og kromofile. Sistnevnte er igjen delt inn i acidofile og basofile (en detaljert histologisk beskrivelse av hypofysen er gitt i den tilsvarende delen av manualen). Det bør imidlertid bemerkes at hormonene som produseres av kjertelcellene som utgjør parenkymet til adenohypofysen, på grunn av mangfoldet av sistnevnte, til en viss grad er forskjellige i sin kjemiske natur, og den fine strukturen til de sekreterende cellene må samsvare med biosynteseegenskapene til hver av dem. Men noen ganger kan overgangsformer av kjertelceller som er i stand til å produsere flere hormoner observeres i adenohypofysen. Det finnes bevis for at typen kjertelceller i adenohypofysen ikke alltid er genetisk bestemt.
Under mellomgulvet til sella turcica ligger den traktformede delen av forlappen. Den omslutter stilken til hypofysen og er i kontakt med den grå tuberkelen. Denne delen av adenohypofysen er preget av tilstedeværelsen av epitelceller og rikelig blodtilførsel. Den er også hormonelt aktiv.
Den mellomliggende (midtre) delen av hypofysen består av flere lag med store sekretorisk aktive basofile celler.
Hypofysen utfører ulike funksjoner gjennom hormonene sine. Den fremre lappen produserer adrenokortikotrope (ACTH), tyreoidstimulerende (TSH), follikkelstimulerende (FSH), luteiniserende (LH), lipotrope hormoner, samt veksthormon - somatotropisk (STO) og prolaktin. I den mellomliggende lappen syntetiseres melanocyttstimulerende hormon (MSH), og i den bakre lappen akkumuleres vasopressin og oksytocin.
ACTH
Hypofysehormoner er en gruppe protein- og peptidhormoner og glykoproteiner. Av hormonene i den fremre hypofysen har ACTH blitt studert best. Det produseres av basofile celler. Dets viktigste fysiologiske funksjon er å stimulere biosyntesen og utskillelsen av steroidhormoner fra binyrebarken. ACTH viser også melanocyttstimulerende og lipotropisk aktivitet. I 1953 ble det isolert i ren form. Senere ble dets kjemiske struktur etablert, bestående av 39 aminosyrerester hos mennesker og en rekke pattedyr. ACTH har ingen artsspesifisitet. For tiden er det utført kjemisk syntese av både selve hormonet og forskjellige fragmenter av molekylet, mer aktive enn naturlige hormoner. Hormonets struktur har to seksjoner av peptidkjeden, hvorav den ene sikrer deteksjon og binding av ACTH til reseptoren, og den andre gir en biologisk effekt. Det binder seg tilsynelatende til ACTH-reseptoren gjennom samspillet mellom elektriske ladninger i hormonet og reseptoren. Rollen til den biologiske effektoren av ACTH utføres av fragmentet av 4-10-molekylet (Met-Glu-His-Phen-Arg-Tri-Tri).
Den melanocyttstimulerende aktiviteten til ACTH skyldes tilstedeværelsen i molekylet av den N-terminale regionen, som består av 13 aminosyrerester og gjentar strukturen til alfa-melanocyttstimulerende hormon. Denne regionen inneholder også et heptapeptid som finnes i andre hypofysehormoner og har noen adrenokortikotrope, melanocyttstimulerende og lipotrope aktiviteter.
Det viktigste øyeblikket i ACTHs virkning anses å være aktiveringen av enzymet proteinkinase i cytoplasmaet med deltakelse av cAMP. Fosforylert proteinkinase aktiverer enzymet esterase, som omdanner kolesterolestere til et fritt stoff i fettdråper. Proteinet som syntetiseres i cytoplasmaet som et resultat av fosforylering av ribosomer stimulerer bindingen av fritt kolesterol til cytokrom P-450 og overføringen av det fra lipiddråper til mitokondrier, hvor alle enzymene som sikrer omdannelsen av kolesterol til kortikosteroider er tilstede.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Skjoldbruskstimulerende hormon
TSH - tyrotropin - hovedregulatoren for utvikling og funksjon av skjoldbruskkjertelen, prosessene for syntese og utskillelse av skjoldbruskkjertelhormoner. Dette komplekse proteinet - glykoprotein - består av alfa- og beta-underenheter. Strukturen til den første underenheten sammenfaller med alfa-underenheten til luteiniserende hormon. Dessuten sammenfaller den i stor grad hos forskjellige dyrearter. Sekvensen av aminosyrerester i beta-underenheten til menneskelig TSH er dechiffrert og består av 119 aminosyrerester. Det kan bemerkes at beta-underenhetene til menneskelig og storfe TSH er like på mange måter. De biologiske egenskapene og naturen til den biologiske aktiviteten til glykoproteinhormoner bestemmes av beta-underenheten. Den sikrer også hormonets interaksjon med reseptorer i forskjellige målorganer. Imidlertid viser beta-underenheten hos de fleste dyr spesifikk aktivitet først etter at den kombineres med alfa-underenheten, som fungerer som en slags aktivator av hormonet. Sistnevnte induserer med like stor sannsynlighet luteiniserende, follikkelstimulerende og tyrotropiske aktiviteter bestemt av egenskapene til beta-underenheten. Likheten som er funnet lar oss konkludere med at disse hormonene oppsto i evolusjonsprosessen fra en felles forgjenger, og beta-underenheten bestemmer også hormonenes immunologiske egenskaper. Det er en antagelse om at alfa-underenheten beskytter beta-underenheten mot virkningen av proteolytiske enzymer, og også letter transporten fra hypofysen til de perifere målorganene.
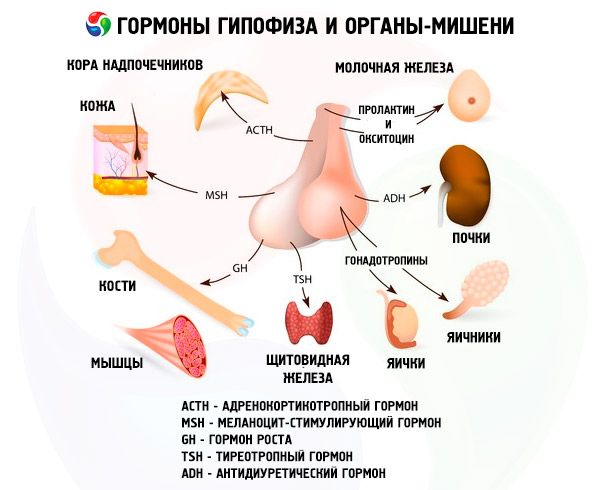
Gonadotrope hormoner
Gonadotropiner finnes i kroppen i form av LH og FSH. Det funksjonelle formålet med disse hormonene er generelt redusert til å sikre reproduksjonsprosesser hos individer av begge kjønn. De er, i likhet med TSH, komplekse proteiner - glykoproteiner. FSH induserer modning av follikler i eggstokkene hos kvinner og stimulerer spermatogenesen hos menn. LH forårsaker follikkelruptur hos kvinner med dannelse av corpus luteum og stimulerer utskillelsen av østrogener og progesteron. Hos menn akselererer det samme hormonet utviklingen av interstitielt vev og utskillelsen av androgener. Effektene av gonadotropiner er avhengige av hverandre og forekommer synkront.
Dynamikken i gonadotropinsekresjonen hos kvinner endrer seg i løpet av menstruasjonssyklusen og har blitt studert i tilstrekkelig detalj. I den preovulatoriske (follikulære) fasen av syklusen er LH-innholdet på et ganske lavt nivå, mens FSH økes. Etter hvert som follikkelen modnes, øker østradiolsekresjonen, noe som fremmer økt produksjon av gonadotropiner fra hypofysen og forekomsten av både LH- og FSH-sykluser, dvs. kjønnssteroider stimulerer utskillelsen av gonadotropiner.
For tiden er strukturen til LH bestemt. I likhet med TSH består den av to underenheter: a og b. Strukturen til alfa-underenheten til LH hos forskjellige dyrearter samsvarer i stor grad med strukturen til alfa-underenheten til TSH.
Strukturen til beta-underenheten til LH er betydelig forskjellig fra strukturen til beta-underenheten til TSH, selv om den har fire identiske seksjoner av peptidkjeden som består av 4–5 aminosyrerester. I TSH er de lokalisert i posisjonene 27–31, 51–54, 65–68 og 78–83. Siden beta-underenheten til LH og TSH bestemmer den spesifikke biologiske aktiviteten til hormonene, kan det antas at homologe seksjoner i strukturen til LH og TSH bør sikre forbindelsen mellom beta-underenheter og alfa-underenheten, og seksjoner som er forskjellige i struktur, bør være ansvarlige for spesifisiteten til hormonenes biologiske aktivitet.
Naturlig LH er svært stabilt mot virkningen av proteolytiske enzymer, men beta-subenheten spaltes raskt av chymotrypsin, og a-subenheten er vanskelig å hydrolysere av enzymet, dvs. den spiller en beskyttende rolle og hindrer chymotrypsin i å få tilgang til peptidbindingene.
Når det gjelder den kjemiske strukturen til FSH, har forskerne ennå ikke oppnådd definitive resultater. I likhet med LH består FSH av to underenheter, men beta-underenheten til FSH er forskjellig fra beta-underenheten til LH.
Prolaktin
Et annet hormon, prolaktin (laktogent hormon), spiller en aktiv rolle i reproduksjonsprosesser. De viktigste fysiologiske egenskapene til prolaktin hos pattedyr manifesterer seg i form av stimulering av utviklingen av melkekjertler og laktasjon, vekst av talgkjertler og indre organer. Det fremmer manifestasjonen av effekten av steroider på sekundære seksuelle egenskaper hos hanner, stimulerer den sekretoriske aktiviteten til corpus luteum hos mus og rotter, og deltar i reguleringen av fettmetabolismen. Mye oppmerksomhet har blitt viet prolaktin de siste årene som en regulator av mors atferd; slik polyfunksjonalitet forklares av dets evolusjonære utvikling. Det er et av de gamle hypofysehormonene og finnes til og med hos amfibier. For tiden har strukturen til prolaktin hos noen pattedyrarter blitt fullstendig dechiffrert. Inntil nylig uttrykte forskere imidlertid tvil om eksistensen av et slikt hormon hos mennesker. Mange trodde at dets funksjon ble utført av veksthormonet. Nå er det innhentet overbevisende bevis for tilstedeværelsen av prolaktin hos mennesker, og strukturen er delvis dechiffrert. Prolaktinreseptorer binder seg aktivt til veksthormon og placentalaktogen, noe som indikerer en enkelt virkningsmekanisme for de tre hormonene.
Somatotropin
Veksthormonet somatotropin har et enda bredere virkningsspekter enn prolaktin. I likhet med prolaktin produseres det av acidofile celler i adenohypofysen. STH stimulerer skjelettvekst, aktiverer proteinbiosyntese, har en fettmobiliserende effekt og fremmer en økning i kroppsstørrelse. I tillegg koordinerer det metabolske prosesser.
Hormonets deltakelse i sistnevnte bekreftes av en kraftig økning i utskillelsen av hypofysen, for eksempel når blodsukkernivået synker.
Den kjemiske strukturen til dette menneskelige hormonet er nå fullt etablert – 191 aminosyrerester. Primærstrukturen ligner på korionisk somatomammotropin eller placentalaktogen. Disse dataene indikerer en betydelig evolusjonær nærhet mellom de to hormonene, selv om de viser forskjeller i biologisk aktivitet.
Det er nødvendig å understreke den høye artsspesifisiteten til det aktuelle hormonet - for eksempel er STH av animalsk opprinnelse inaktivt hos mennesker. Dette forklares både av reaksjonen mellom reseptorene til STH hos mennesker og dyr, og selve hormonets struktur. For tiden pågår studier for å identifisere aktive sentre i den komplekse strukturen til STH som viser biologisk aktivitet. Individuelle fragmenter av molekylet som viser andre egenskaper studeres. For eksempel, etter hydrolyse av humant STH med pepsin, ble et peptid bestående av 14 aminosyrerester og tilsvarende molekylseksjonen 31-44 isolert. Det hadde ingen veksteffekt, men overgikk det native hormonet betydelig i lipotropisk aktivitet. Humant veksthormon, i motsetning til lignende hormoner hos dyr, har betydelig laktogen aktivitet.
Adenohypofysen syntetiserer mange peptid- og proteinstoffer som har en fettmobiliserende effekt, og de hypofysetropiske hormonene - ACTH, STH, TSH og andre - har en lipotropisk effekt. I de senere år har beta- og y-lipotropiske hormoner (LPG) blitt spesielt vektlagt. De biologiske egenskapene til beta-LPG har blitt studert i detalj; i tillegg til lipotropisk aktivitet har det også en melanocyttstimulerende, kortikotropinstimulerende og hypokalsemisk effekt, og gir også en insulinlignende effekt.
For tiden er den primære strukturen til LPG hos sau (90 aminosyrerester), lipotropiske hormoner hos griser og storfe, dechiffrert. Dette hormonet har artsspesifisitet, selv om strukturen til den sentrale regionen av beta-LPG er den samme i forskjellige arter. Det bestemmer hormonets biologiske egenskaper. Et av fragmentene av denne regionen finnes i strukturen til alfa-MSH, beta-MSH, ACTH og beta-LPG. Det antydes at disse hormonene oppsto fra den samme forløperen i evolusjonsprosessen. γ-LPG har en svakere lipotropisk aktivitet enn beta-LPG.
Melanocyttstimulerende hormon
Dette hormonet, syntetisert i den mellomliggende lappen i hypofysen, stimulerer biosyntesen av hudpigmentet melanin i dets biologiske funksjon, fremmer en økning i størrelsen og antallet pigmentceller melanocytter i huden hos amfibier. Disse egenskapene til MSH brukes i biologisk testing av hormonet. Det finnes to typer hormon: alfa- og beta-MSH. Det har blitt vist at alfa-MSH ikke har artsspesifisitet og har samme kjemiske struktur hos alle pattedyr. Molekylet er en peptidkjede som består av 13 aminosyrerester. Beta-MSH har derimot artsspesifisitet, og strukturen varierer hos forskjellige dyr. Hos de fleste pattedyr består beta-MSH-molekylet av 18 aminosyrerester, og bare hos mennesker er det forlenget fra aminosyreenden med fire aminosyrerester. Det skal bemerkes at alfa-MSH har en viss adrenokortikotropisk aktivitet, og effekten på dyrs og menneskers atferd er nå bevist.
Oksytocin og vasopressin
Vasopressin og oksytocin, som syntetiseres i hypothalamus, akkumuleres i hypofysens bakre lobb: vasopressin i nevronene i nucleus supraopticus, og oksytocin i nucleus paraventrikulatoires. Deretter overføres de til hypofysen. Det bør understrekes at forløperen til vasopressinhormonet syntetiseres først i hypothalamus. Samtidig produseres proteinet nevrofysin type 1 og 2 der. Det første binder seg til oksytocin, og det andre binder seg til vasopressin. Disse kompleksene migrerer i form av nevrosekretoriske granuler i cytoplasmaet langs aksonet og når hypofysens bakre lobb, hvor nervefibrene ender i karveggen og innholdet i granulene kommer inn i blodet. Vasopressin og oksytocin er de første hypofysehormonene med en fullstendig etablert aminosyresekvens. I sin kjemiske struktur er de nonapeptider med én disulfidbro.
Hormonene som vurderes har ulike biologiske effekter: de stimulerer transporten av vann og salter gjennom membraner, har en vasopressoreffekt, forsterker sammentrekninger av livmorens glatte muskulatur under fødsel og øker utskillelsen fra melkekjertlene. Det bør bemerkes at vasopressin har en høyere antidiuretisk aktivitet enn oksytocin, mens sistnevnte har en sterkere effekt på livmoren og melkekjertlene. Hovedregulatoren for vasopressinsekresjon er vannforbruk; i nyretubuli binder det seg til reseptorer i de cytoplasmatiske membranene med påfølgende aktivering av enzymet adenylatcyklase i dem. Ulike deler av molekylet er ansvarlige for å binde hormonet til reseptoren og for den biologiske effekten.
Hypofysen, som er forbundet med hele nervesystemet gjennom hypothalamus, forener det endokrine systemet til en funksjonell helhet og bidrar til å sikre at kroppens indre miljø holder seg stabilt (homeostase). Innenfor det endokrine systemet utføres homeostatisk regulering basert på prinsippet om tilbakekobling mellom hypofysens fremre lob og "målkjertlene" (skjoldbruskkjertelen, binyrebarken, gonader). Overskudd av hormonet som produseres av "målkjertelen" hemmer, og mangel på det stimulerer utskillelsen og frigjøringen av det tilsvarende tropiske hormonet. Hypothalamus er en del av tilbakekoblingssystemet. Det er i det reseptorsonene som er følsomme for hormonene i "målkjertlene" befinner seg. Ved å binde seg spesifikt til hormoner som sirkulerer i blodet og endre responsen avhengig av hormonkonsentrasjonen, overfører hypothalamiske reseptorer sin effekt til de tilsvarende hypothalamiske sentrene, som koordinerer arbeidet til adenohypofysen og frigjør hypothalamiske adenohypofysiotrope hormoner. Dermed bør hypothalamus betraktes som en nevroendokrin hjerne.