Medisinsk ekspert av artikkelen

Nye publikasjoner
Escherichioser (slekten Escherichia, E. coli)
Sist anmeldt: 06.07.2025

Alt iLive-innhold blir gjennomgått med medisin eller faktisk kontrollert for å sikre så mye faktuell nøyaktighet som mulig.
Vi har strenge retningslinjer for innkjøp og kun kobling til anerkjente medieområder, akademiske forskningsinstitusjoner og, når det er mulig, medisinsk peer-evaluerte studier. Merk at tallene i parenteser ([1], [2], etc.) er klikkbare koblinger til disse studiene.
Hvis du føler at noe av innholdet vårt er unøyaktig, utdatert eller ellers tvilsomt, velg det og trykk Ctrl + Enter.
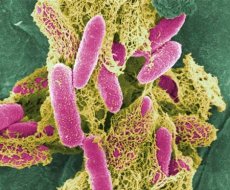
Hovedrepresentanten for slekten Escherichia - E. coli - ble først oppdaget i 1885 av T. Escherich, etter hvem denne bakterieslekten fikk navnet sitt. Viktige trekk ved denne slekten: peritrik (eller ikke-bevegelig), fermenterer laktose med dannelse av syre og gass (eller laktose-negativ), vokser ikke på et sultemedium med sitrat, Voges-Proskauer-reaksjonen er negativ, MR-testen er positiv, har ikke fenylalanin-deaminase, vokser ikke på et medium med KCN, innholdet av G + C i DNA er 50-51 mol%.
Slekten Escherichia omfatter minst 7 arter; av spesiell betydning innen medisin er arten E. coli, spesielt de variantene som forårsaker menneskelige sykdommer. De er delt inn i to hovedgrupper: de som forårsaker ekstraintestinale sykdommer og de som forårsaker akutte tarmsykdommer (AIDS). Representanter for den første er delt inn i tre patologiske grupper:
- meningeal (MENEC - meningitt E. coli);
- septikemi (SEPEC - septikemi E. coli) og
- uropatogen (UPEC - uropatogen E. coli).
E. coli-variantene som forårsaker akutte luftveisinfeksjoner ble i sin tur først delt inn i følgende fire kategorier: enterotoksigene E. coli (ETEC); enteroinvasive E. coli (EIEC); enteropatogen E. coli (EPEC) og enterohemorragisk E. coli (EHEC). Deretter ble to kategorier identifisert: enteroaggregativ E. coli (EAEC) og diffust aggregerende E. coli (DAEC).
I tillegg brukes E. coli i internasjonale standarder som en indikator på graden av fekal forurensning av vann, spesielt drikkevann og matvarer.
Standardstammen av E. coli (E. coli K-12) er mye brukt i laboratorier i mange land rundt om i verden for å studere bakterienes genetikk.
 [
[ Morfologi
E. coli er en fakultativ anaerob bakterie som vokser godt på konvensjonelle næringsmedier – kolonier på agar er runde, konvekse og gjennomskinnelige. Veksten på buljong skjer i form av diffus turbiditet. Temperaturoptimum for vekst er 37 °C, vokser i området fra 10 til 45 °C, og optimal pH er 7,2–7,5. På alle differensialdiagnostiske medier er kolonier av E. coli som dekomponerer laktose farget i indikatorens farge (på Endo-medium – mørk karmosinrød med metallisk glans).
Biokjemiske egenskaper
I de fleste tilfeller er E. coli i stand til å fermentere følgende karbohydrater for å danne syre og gass: glukose, laktose, mannitol, arabinose, galaktose, noen ganger sukrose og noen andre karbohydrater; danner indol; danner vanligvis ikke H2S ; reduserer nitrater til nitritter, flyter ikke gelatin, vokser ikke på et sultemedium med sitrat, gir en positiv reaksjon med MR og en negativ reaksjon med Voges-Proskauer. Ved disse tegnene kan den lett skilles fra patogener av en rekke sykdommer (dysenteri, tyfusfeber, salmonellose, etc.). Patogene E. coli skiller seg imidlertid ofte ikke fra ikke-patogene, verken ved kulturelle eller biokjemiske egenskaper.
Patogenitetsfaktorer for E. coli
E. colis evne til å forårsake ulike sykdommer skyldes tilstedeværelsen av følgende patogenitetsfaktorer:
Adhesjons- og koloniseringsfaktorer. De er nødvendige for feste til vevsceller og deres kolonisering. Tre varianter av koloniseringsfaktoren er oppdaget: a) CFA/I-CFA/VI (koloniseringsfaktor) - de har en fimbrial struktur; b) EAF (enteropatogen E. coli adhesjonsfaktor) - intimin - et ytre membranprotein, kodet av eaeA-genet. Funnet i 4 og EHEC, detekteres det ved bakterienes evne til å feste seg til Hep-2-celler; c) Adhesjon Henle-407 - fimbrialstrukturer, detekteres ved bakterienes evne til å feste seg til Henle-407-celler. Alle er kodet av plasmidgener. I tillegg til dem er andre koloniseringsfaktorer beskrevet, som også kan inkludere bakterielle lipopolysakkarider.
Invasjonsfaktorer. Med deres hjelp trenger for eksempel EIEC og EHEC inn i tarmepitelceller, formerer seg i dem og forårsaker deres ødeleggelse. Rollen til invasjonsfaktorene utføres av proteiner i den ytre membranen.
Eksotoksiner. Det er funnet at patogene E. coli inneholder eksotoksiner som skader membraner (hemolysin), hemmer proteinsyntese (Shiga-toksin) og aktiverer sekundære budbringere (budbringer - kommunikasjon) - toksiner CNF, ST, CT, CLTD, EAST.
Hemolysiner produseres av forskjellige patogener, inkludert E. coli. Hemolysin er et poredannende toksin. Det binder seg først til målcellemembranen og danner deretter en pore i den, gjennom hvilken små molekyler og ioner kommer inn og ut, noe som fører til celledød og erytrocyttlyse.
Shigatoksin (STX) ble først oppdaget i Shigella dysenteriae, og deretter ble et lignende toksin (Shiga-lignende toksin) funnet i EHEC. Toksinet (N-glykosidase) blokkerer proteinsyntesen ved å samhandle med 28S rRNA, noe som resulterer i celledød (cytotoksin). Det finnes to typer Shiga-lignende toksiner: STX-1 og STX-2. STX-1 er nesten identisk med Shigatoksin i sine antigene egenskaper, mens STX-2 er forskjellig fra Shigatoksin i sine antigene egenskaper, og i motsetning til STX-1 nøytraliseres det ikke av antiserum mot det. Syntesen av STX-1- og STX-2-cytotoksiner kontrolleres i E. coli av genene til de moderat konverterende profagene 9331 (STX-1) og 933W (STX-2).
- Toksin L (varmelabilt toksin) er en ADP-ribosyltransferase; ved å binde seg til et G-protein forårsaker det diaré.
- ST-toksin (termostabilt toksin), som interagerer med guanylatcyklasereseptoren, stimulerer aktiviteten og forårsaker diaré.
- CNF (cytotoksisk nekrotisk faktor) er et deamidaseprotein som skader såkalte RhoG-proteiner. Dette toksinet finnes i UPEC, som forårsaker urinveisinfeksjoner.
- CLTD-toksin er et cytoletalt, disintegrerende toksin. Virkningsmekanismen er dårlig forstått.
- EAST-toksin er et varmestabilt toksin fra enteroaggregativ E. coli (EAEC), sannsynligvis likt varmestabilt toksin (ST).
Endotoksiner er lipopolysakkarider. De bestemmer bakterienes antigene spesifisitet (som bestemmes av den repeterende sidekjeden av sukkerarter) og formen på koloniene (tap av sidekjeder fører til transformasjon av S-kolonier til R-kolonier).
Dermed kontrolleres patogenitetsfaktorene til E. coli ikke bare av kromosomgener i vertscellen, men også av gener introdusert av plasmider eller tempererte konverterende fager. Alt dette indikerer muligheten for fremvekst av patogene varianter av E. coli som et resultat av spredning av plasmider og tempererte fager blant dem. Nedenfor er en kort beskrivelse av 4 kategorier av E. coli som forårsaker akutte luftveisinfeksjoner; informasjon om de nylig identifiserte kategoriene DAEC og EAEC ble ikke funnet i kildene som er tilgjengelige for oss.
ETEC inkluderer 17 serogrupper. Adhesjons- og koloniseringsfaktorene til fimbriestrukturen av CFA-typen og enterotoksiner (LT eller ST, eller begge) er kodet av samme plasmid(er). De koloniserer villi uten å skade dem. Enterotoksiner forårsaker et brudd på vann-salt-metabolismen. Lokaliseringen av prosessen er tynntarmen. Den infeksiøse dosen er 108–1010 celler. Sykdommen utvikler seg som koleralignende diaré. Typen epidemier er vannbåren, sjeldnere matbåren. Barn i alderen 1 til 3 år og voksne er rammet.
EIEC inkluderer 9 serogrupper, patogenitet er assosiert med evnen til å trenge inn i epitelcellene i tarmslimhinnen og formere seg inni dem, noe som forårsaker deres ødeleggelse. Disse egenskapene er kodet, i tillegg til kromosomale gener, av plasmidgener (140 MD). Plasmidet koder for syntesen av proteiner i den ytre membranen, som bestemmer invasjonen. Både selve plasmidet og proteinene det koder for er relatert til proteinene til dysenteripatogener, noe som forklarer likheten mellom EIEC og shigella. Den smittsomme dosen er 10s celler. Lokaliseringen av prosessen er nedre ileum og tykktarmen. Sykdommen utvikler seg som dysenteri: først vannaktig diaré, deretter kolittsyndrom. Barn 1,5-2 år, ungdom og voksne er syke. Type utbrudd - mat, vann.
Epidemiologi
E. coli er en representant for den normale mikrofloraen i tarmkanalen hos alle pattedyr, fugler, reptiler og fisk. For å avklare spørsmålet om hvilke varianter av E. coli og hvorfor de forårsaker escherichiose, var det derfor nødvendig å studere antigenstrukturen, utvikle en serologisk klassifisering som er nødvendig for å identifisere patogene serovarianter, og finne ut hvilke patogenitetsfaktorer de har, dvs. hvorfor de er i stand til å forårsake ulike former for escherichiose.
E. coli har 171 O-antigenvarianter (01-0171), 57 H-antigenvarianter (H1-H57) og 90 overflate- (kapsel-) K-antigenvarianter. I realiteten finnes det imidlertid 164 O-antigengrupper og 55 H-antigen-serovarianter, siden noen av de tidligere 0:H-serogruppene ble ekskludert fra E. coli-artene, men ordinalnumrene til O- og H-antigenene forble uendret. De antigene egenskapene til diaréfremkallende E. coli inkluderer O- og H-antigennumrene, for eksempel 055:116; 0157:H7; O-antigen indikerer tilhørighet til en bestemt serogruppe, og H-antigen er dens serovariant. I tillegg avdekket en mer grundig studie av O- og H-antigener de såkalte faktor O- og H-antigener, dvs. deres antigene subvarianter, som H2a, H2b, H2c eller O20, O20a, O20ab, etc. Totalt inkluderer listen over diaréfremkallende E. coli 43 O-serogrupper og 57 OH-serovarianter. Denne listen suppleres stadig med nye serovarianter.
Symptomer
Gruppen inkluderer 9 klasse 1 serogrupper og fire klasse 2 serogrupper. Klasse 1 serogrupper har et plasmid (60 MDa) som kontrollerer syntesen av adhesjons- og koloniseringsfaktoren av EAF-typen. Det er representert av et protein lokalisert i den ytre membranen og detekteres av bakterienes evne til å feste seg til HEp-2-celler. Proteinet har en mw på 94 kDa. Klasse 2 serogrupper har ikke dette plasmidet; deres patogenitet skyldes noen andre faktorer. Noen stammer av begge klasser har vist seg å være i stand til å syntetisere STX.4 koloniserer plasmamembranen til enterocytter, noe som forårsaker skade på epiteloverflaten med dannelse av erosjoner og moderat betennelse. Den smittsomme dosen er 105-10 12 celler. Prosessen er lokalisert i tynntarmen. Sykdommen er preget av vannaktig diaré og alvorlig dehydrering. For det meste er barn i det første leveåret syke. Smittemetoden er kontakt-husholdning, sjeldnere matbåren.
Serogruppene EIEC og 4 er de vanligste årsakene til nosokomiale utbrudd.
EHEC-er produserer cytotoksinene STX-1 og STX-2. De forårsaker hemoragisk kolitt hos mennesker med alvorlige komplikasjoner som hemolytisk uremi og trombotisk trombocytopenisk purpura. Toksinene ødelegger endotelcellene i små blodårer. Dannelsen av blodpropper og fibrinavsetning fører til nedsatt blodstrøm, blødning, iskemi og nekrose i celleveggen. Uremisk hemolytisk syndrom kan være dødelig. EHEC-er er representert av mange serotyper (-150), men den viktigste epidemiologiske rollen spilles av E. coli 0157-H7 og dens flagellatmutant E. coli 0157:NM, siden bare de produserer STX. Disse bakteriestammene kan bare skille ut ett av cytotoksinene eller begge samtidig. Det antas at det naturlige reservoaret for EHEC-serovarer, inkludert E. coli 0157:H7, er storfe og sau. Den vanligste smitteveien er mat (kjøtt, spesielt kjøttdeig; melk). E. coli 0157:H7 er uvanlig resistent mot ugunstige faktorer. Dette bidrar til dens overlevelse og reproduksjon i ulike produkter. Infeksjon er mulig gjennom kontakt med husholdningsartikler. Sykdommen starter akutt: tarmkramper oppstår, deretter diaré, først vannaktig, deretter blodig. Barn og voksne er syke. En syk person er smittsom.
Laboratoriediagnostikk
Den er basert på isolering av en renkultur av patogenet og dens identifisering, samt testing av toksiner ved hjelp av PCR. Det forårsakende agenset til escherichiose identifiseres ved hjelp av et sett med polyvalente OK-sera og et sett med adsorberte sera som kun inneholder antistoffer mot visse antigener. En keratokonjunktivaltest kan brukes til å identifisere EIEC. Noen representanter for EIEC er immobile og fermenterer ikke laktose og salicin. Identifisering av E. coli 0157:H7 forenkles av dens manglende evne til å fermentere sorbitol (endomedium med sorbitol brukes i stedet for laktose). Men det er best å bruke PCR-testsystemer for å identifisere og differensiere OKZ-patogener (alle kategorier). Om nødvendig bestemmes følsomheten til de isolerte patogenene for antibiotika.